Рис. 14.4(epig). Эпигенетическая регуляция функций теломер
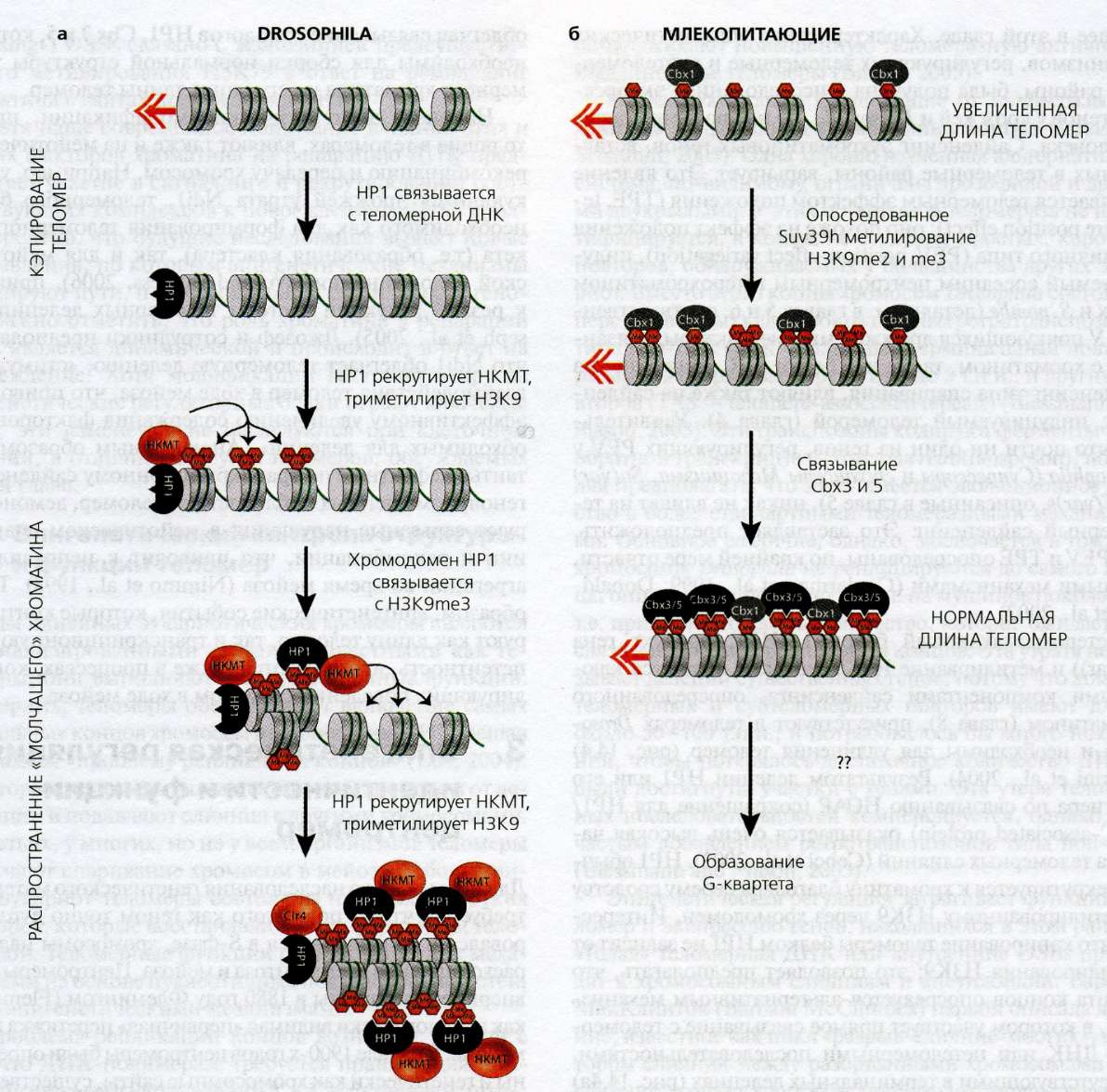
У мух и млекопитающих функции теломер регулируются эпигенетически.
(а) У Drosophila гетерохроматиновый белок 1 (НР1) связывается с теломерной ДНК независимо от ее хромодомена и "кэпирует" теломеры, что обеспечивает нормальное расхождение благодяря блокировке теломерных слияний ( Fanti et al. 1998 ; Perrini et al. 2004 ). Затем HP1 рекрутирует неизвестную метилтрансферазу гистонов (НКМТ; не Su(var)3-9), которая триметилирует НЗК9 на близлежащих нуклеосомах; НР1 связывается с НЗК9mеЗ через хромодомен, что, в свою очередь, рекрутирует большие количества НКМТ, а последовательные раунды связывания НР1 - рекрутирования НКМТ стимулируют распространение "молчащего" хроматина по субтеломерным районам.
(б) У мышей нокаут обоих локусов НКМТ Suv39 понижает уровни НЗК9mеЗ и mе2 и увеличивает содержание модификаций НЗК9mе, измененной структуры хроматина и изменений в уровнях белков, связывающихся с ди- и триметилированными НЗК9 (стрелка вниз Cbx 3 и 5), H3K9me (стрелка вверх Cbx 1) и TERF1 и 2 (не показано) в теломерах ( Garcia-Cao et al. 2004 ). Эти изменения коррелируют с увеличенной длиной теломер, позволяя предполагать, что триметилирование НЗК9 с помощью Suv39hs необходимо для нормального функционирования теломеразы и регуляции размера теломер.