Рис. 3.20(epig). Эпигенетическое усиление первичного сигнала
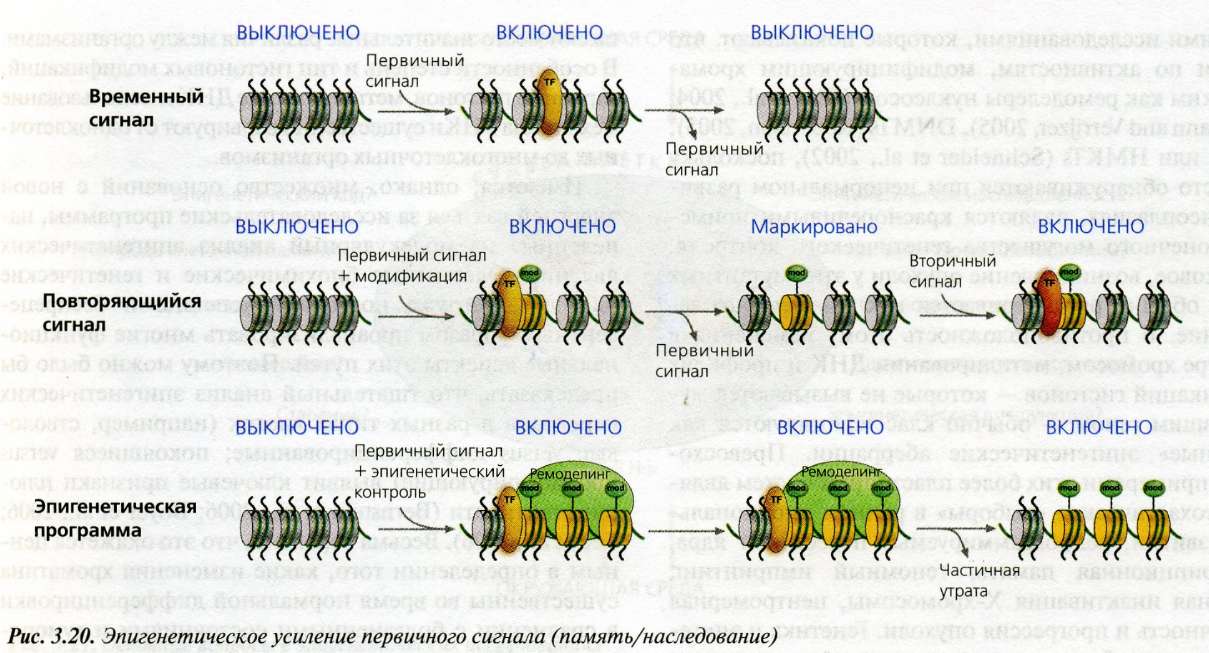
Классическая генетика предсказывает, что экспрессия гена зависит от наличия и связывания соответствующего набора транскрипционных факторов (TF). Удаление таких факторов (т.е. первичный сигнал) приводит к утрате экспрессии гена и, таким образом, составляет преходящий активирующий сигнал (верхняя часть рисунка). Структура хроматина вносит свой вклад в экспрессию гена: здесь некоторые конформации являются репрессивными, а другие активными. Активация локуса может поэтому происходить посредством первичного сигнала и приводить к изменению в структуре хроматина "вниз по течению", включающему активные ковалентные гистоновые метки (mod) и замещение коровых гистонов их вариантами (например, НЗ.З). При клеточном делении эта структура хроматина может быть восстановлена в присутствии активирующего сигнала (обозначается как "повторяющийся сигнал"). Результатом эпигенетической памяти оказывается поддержание состояния хроматина при клеточном делении, даже в отсутствие первичного активирующего сигнала. Такая система памяти не является абсолютной, но включает множественные уровни эпигенетического регулирования для ремоделинга структуры хроматина.
Динамическая природа хроматина означает, что хотя состояние хроматина может быть митотически стабильным, оно тем не менее склонно к изменению, влияя тем самым на продолжительность эпигенетической памяти.