Рис. 4.3(epig). Переключение типов спаривания у дрожжей
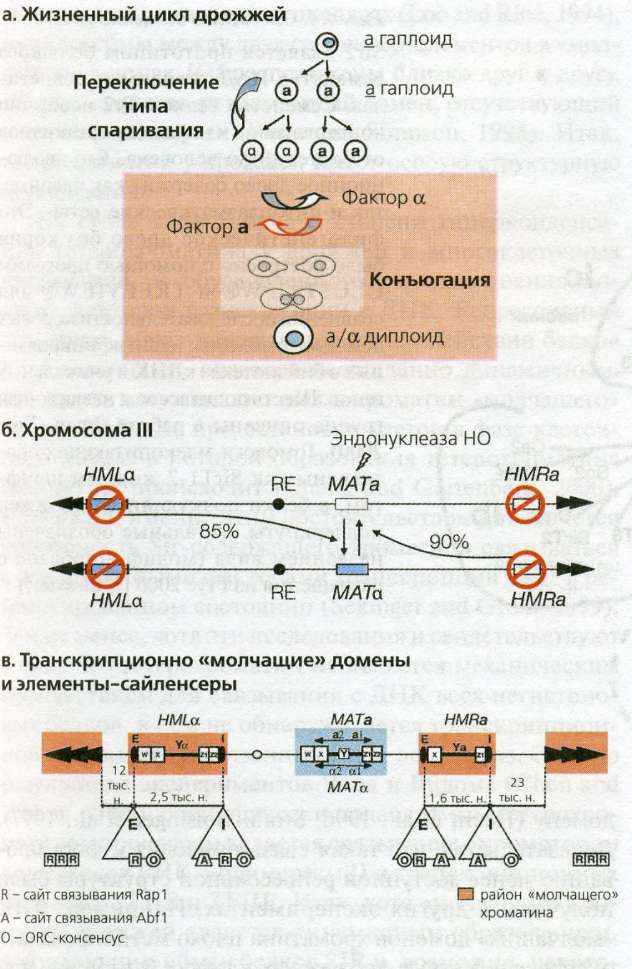
(а) Гомоталлические штаммы дрожжей способны переключать тип спаривания с каждым клеточным циклом. Переключение происходит до репликации ДНК, поэтому и материнская, и дочерняя клетки приобретают новый тип спаривания.
(б) Показано положение "молчащего" и экспрессирующегося локусов типа спаривания на III хромосоме. Активный локус MAT может переключаться посредством конверсии генов примерно один раз на каждый клеточный цикл, благодаря двунитчатым разрывам ДНК эндонуклеазой НО. Указанные проценты показывают частоту, с которой события конверсии переключают локусы МАТ между противоположными типами. Направленность переключения обеспечивается энхансером рекомбинации (RE) на левом плече III хромосомы.
(в) Репрессия "молчащих" локусов типов спаривания HMR и HML происходит под действием двух сайленсеров, фланкирующих эти гены. Они называются Е (от essential) и I (от important) ( Brand et al., 1987 ) и содержат участки связывания белков Rap1 (R), Abf1 (A) и ORC (О). Комбинируя такие сайты связывания в разных комбинациях, можно создавать искусственные сайленсеры, хотя их эффективность ниже, чем у природных. HMLaльфа и HMRa расположены на расстоянии 12 и 23 т.п.н. от теломеры III хромосомы, соответственно. Домены теломерного гетерохроматина III хромосомы репрессируются независимо от локусов НМ процессом, инициируемым множественными сайтами связывания Rap1 (R).