Повторы ДНК и мутации
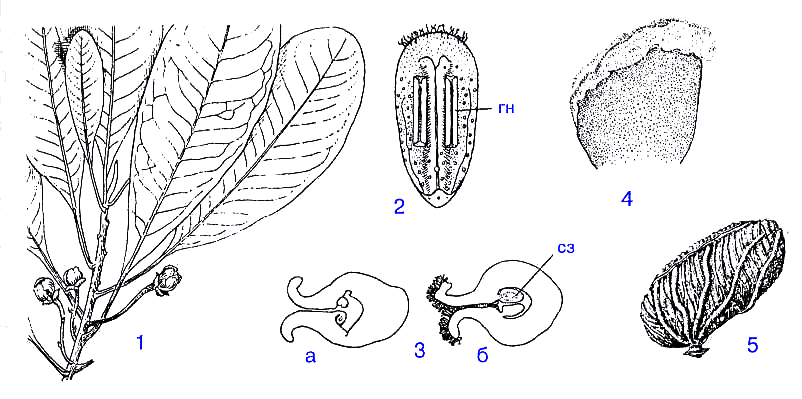
Повторы генерируют не только чреватые прогрессивными нововведениями дупликации, но и фактически весь известный нам сегодня спектр мутационных и рекомбинационных изменений. Самые захватывающие на сей счет дискуссии развернулись вокруг вопроса от эволюционной роли "прыгающих" генов - МГЭ. Отметим, что уже на уровне отдельных генов несовершенные повторы, несущие локальные нарушения гомологии, - источник (быть может, главный) точковых мутаций. Например, неправильное спаривание по частично комплементарным участкам ДНК образует несовершенные гетеродуплексы, а последующая репарация некомплементарных пар как раз и приводит к заменам нуклеотидов ( рис. 2а , 2б ). ( Ripley, Glickman, 1983 ). Понятно, что подобная "коррекция" может происходить и в собственно структурной, транслируемой, части гена, и в его регуляторной зоне. Так, поскольку большинсто белков имеют явно выраженные периодические элементы вторичной структуры (alpha-спирали, beta-структуры), то соответствующие гены содержат неслучайные прямые повторы и комплементарные палиндромы, причем именно несовершенные ( Ohno, 1984 ).
Если только повторы действительно являются основным поставщиком наследственных вариаций, тогда, грубо говоря, чем их больше в геноме, тем более последний неустойчив. С другой стороны, равномерное их распределение по геному представляется крайне нерациональным, ибо в этом случае слишком частыми были бы нарушения целостности генов. Так что есть свой резон в том, что все эти мутагенные и рекомбиногенные повторы и сайты сосредоточены, как правило, вне структурных генов. Что же касается квазипереодичности вторичной структуры глобулярных белков, то нельзя забывать, что для образования несовершенных гетеродуплексов требуется определенное состояние хроматина; поэтому в норме внутригенные изменения, обусловленные повторами, происходят довольно редко. Единственное известное исключение у высших организмов - аномально интенсивный процесс соматического гипермутагенеза в "зрелых" V-генах, которые кодируют вариабельные домены иммуноглобулинов ( Tonegawa, 1983 ).
В последнее время часто высказывается точка зрения, что раз столь солидная доля изменений связана с повторами, а они вовсе не случайно распределены в геноме, то, следовательно, налицо изначальная структурная предетерминированность и отсюда направленность наследственной изменчивости. Разумеется в процессах генерирования изменчивости нет полного хаотичного простора, гены меняются не кто во что горазд; давно, начиная с классических работ С. Бензера (1960) на локусе rII фага T4, описаны всевозможные "горячие" точки мутирования и рекомбинирования на многих ярусах - от отдельных генов до генома в целом; наконец, трудно переоценить значение открытия, что точки эти совпадают с местами скопления прямых и инвертированных повторов и самокомплементарных палиндромов. И все-таки предопределенности здесь столько же сколько в банальной истине, что "прежде чем сойти с ума, надо его иметь".
К тому же мутационная коррекция несовершенных повторов неминуемо увеличивает их гомологичность и тем самым снижает мутабильность на будущее. Для тго, чтобы расширить спектр потенциальных мутаций, т.е. опять сделать точку "горячей", требуется, наоборот, нарушить комплементарность спаренных участков.
Таким образом, "предетерминированной" повторами и в этом смысле неслучайной изменчивости должны предшествовать нарушения (надо думать, совершенно случайные) идентичности или комплементарности повторов. Так что те, кто исповедуют молекулярный предетерминизм в эволюции, обязаны, дабы быть последовательными, позаботиться о неслучайном, до известной степени направленном возникновении исходного несовершенства повторов.
Но, пожалуй, наиболее откровенно атавистическая тяга к преформизму проявляет себя в спорах об эволюционной значимости подвижных элементов генома.