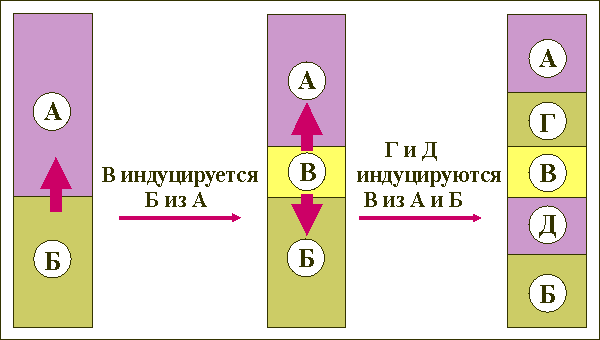
Альфоидная ДНК: трехуровневая организация
Альфа-сателлитная ДНК человека имеет сложную иерархическую организацию и формирует три уровня гомологии [ Willard, Waye, 1987b ].
Первый уровень организации представлен мономером альфоидной ДНК - элементарной, то есть внутренне не повторяющейся, единицей альфа- сателлитной ДНК длиной около 171 н. п. Обычно соседние мономеры ориентированы по типу "голова к хвосту" [ рис. 1 ]. Гомология между последовательностями индивидуальных мономеров и усредненной ( консенсусной) последовательностью всех известных альфа-сателлитных мономеров составляет обычно 60-70%.
Второй уровень организации представлен так называемым предковым повтором - сегментом альфоидной ДНК, содержащим один или несколько мономеров определенных типов, являющимся общим предком для ряда современных хромосомоспецифичных семейств [ Willard, Waye, 1987b ]. Последовательности, имеющие одинаковую структуру предкового повтора, объединяются в одно надхромосомное семейство [ Александров и др., 1986b ; Alexandrov et al., 1988 ; Willard, Waye, 1987a ].
Каждое надхромосомное семейство характеризуется также свойственными только ему типами мономеров . Гомология между консенсусной последовательностью какого-либо типа мономеров и последовательностями индивидуальных мономеров, принадлежащих к этому типу, составляет обычно около 80-90 %.
1 и 2 семейства имеют димерный предковый повтор (типы мономеров этих семейств обозначают как J1-J2 и D1-D2 соответственно [ Alexandrov et al., 1991 ]), 3 семейство характеризуется пентамерной структурой (типы мономеров W1, W2, W3, W4, W5 [ Willard, Waye, 1987a ], обозначения даны по [Alexandrov et al., 1991]), семейство 4 характеризуется мономерной организацией (тип М1), семейство 5 имеет два типа мономеров (R1 и R2), чередующихся неравномерно [ Romanova et al., 1996 ] ( рис 2 каз ).
Последовательности каждого надхромосомного семейства расположены на нескольких хромосомах. Первое надхромосомное семейство локализуется на хромосомах 1, 3, 5, 6, 7, 10, 12, 16 и 19; второе - на хромосомах 2, 4, 8, 9, 13, 14, 15, 18, 20, 21, 22; третье - на хромосомах 1, 11, 17 и Х [ Александров и др., 1986b ; Willard, Waye, 1987a ; Yurov et al., 1987a ; Alexandrov et al., 1988 ].
Первые три надхромосомных семейства охватывают все хромосомы человека, кроме Y и, за исключением хромосом 1 и 18, встречаются только один раз на каждой хромосоме. Локализация надхромосомных семейств 4 и 5 изучена недостаточно.
Предковые мономерные типы не в одинаковой степени гомологичны друг другу. Их можно разделить на две группы:
тип А: J1, D2, W4, W5, M1, R2
тип В: J2, D1, W1, W2, W3, R1 В работе Romanova et al. (1996) показано, что из семи различий между консенсусными последовательностями типов А и В, пять расположены в участке длиной 16 нуклеотидов, представляющем собой сайт связывания белка pJ? [ Gaff et al., 1994 ] (тип А) или сайт связывания белка СЕNР-В (тип В).
В пределах каждого типа сайт связывания СЕNР-В/pJ? представляет собой высоко консервативный участок. Показано, что сайты связывания СЕNР-В существуют у человека и высших приматов, но не у низших приматов, тогда как сайты связывания pJ? широко распространены у низших и высших приматов.
У низших приматов обнаружены только мономеры типа А, в отличие от высших приматов, имеющих мономеры типов А и В. Это свидетельствует о том, что тип А является более древним в эволюционном отношении. Интересно, что консенсусные последовательности типов А и В практически совпадают с последовательностями мономеров R1 и R2. Таким образом, мономеры надхромосомного семейства 5 представляют собой, по-видимому, реликтовые последовательности для типов А и В.
Третьим уровнем организации альфа-сателлитной ДНК является повторяющаяся единица высокого порядка (ПЕВП) , которая определяет уникальность хромосомоспецифичных семейств [ Waye, Willard, 1987 ].
Размер и структура ПЕВП зависят от длины и структуры сегмента, амплифицированного при возникновении данного семейства. Если дивергенция между соседними мономерами составляет приблизительно 10-40%, то дивергенция между повторяющимися единицами высокого порядка, принадлежащими к одному альфа-сателлитному домену, как правило, не превышает 5%. В настоящее время известно несколько десятков хромосомоспецифичных подтипов альфа-сателлитной ДНК, локализованной на всех хромосомах человека [ Lee et al., 1997 ]. Хромосомоспецифичные семейства большинства хромосом не имеют высокогомологичных последовательностей даже в пределах своего надхромосомного семейства, однако некоторые хромосомы могут содержать очень близкие и даже практически не различимые последовательности, например, хромосомы 13 и 21, 14 и 22, 5 и 19 [ Devilee et al., 1986 ; Jorgensen et al., 1988 ; Baldini et al., 1989 ].
Альфа-сателлитные повторы занимают на каждой хромосоме протяженные участки (домены) длиной от 200 тысяч н. п. до 5 миллионов н. п. и более в разных хромосомах, внутри которых ПЕВП повторяются тандемно и практически не различаются по длине и структуре [ Durfy, Willard, 1989 ; Warburton, Willard, 1990 ; Marcais et al., 1991a ]. Трехуровневая организация альфа-сателлитов впервые была показана в экспериментах по гибридизации in situ альфоидных ДНК зондов с хромосомами человека. В условиях низкой жесткости гибридизация наблюдается в прицентромерных районах всех хромосом, что связано с перекрестным узнаванием всех альфоидных мономеров. В условиях средней жесткости альфоидные ДНК зонды гибридизуются только с несколькими хромосомами, содержащими последовательности одного надхромосомного семейства. Наконец, в жестких условиях клонированные альфа-сателлиты гибридизуются обычно только с одной хромосомой, несущей соответствующую данному ДНК зонду повторяющуюся единицу высокого порядка [ Алекандров и др., 1986 ; Юров и др., 1988 ; Alexandrov et al., 1988 ; Yurov et al., 1997 ].
Каждый отдельный структурный уровень в организации последовательностей альфоидной ДНК, по-видимому, является результатом отдельного амплификационного события. При этом "первая" амплификация привела к образованию альфоидного мономера, а последующие амплификационные события привели к появлению различных типов предковых повторов. В свою очередь, каждое хромосомоспецифичное семейство, по-видимому, также возникло в результате отдельного амплификационного события. При этом повторяющаяся единица высокого порядка (ПЕВП, амплификационная единица) представляет собой сегмент альфа-сателлитной последовательности, подвергшийся последней амплификации [ Alexandrov et al., 1988 ]. Следует отметить, что этот механизм эволюции не характерен для надхромосомных семейств 4 и 5 .
Эволюция альфа-сателлитных повторов надхромосомных семейств 4 и 5 не привела к формированию у них повторяющихся единиц высокого порядка.