Рис. 14.7(epig). Паттерны модификаций гистонов в центромерном хроматине
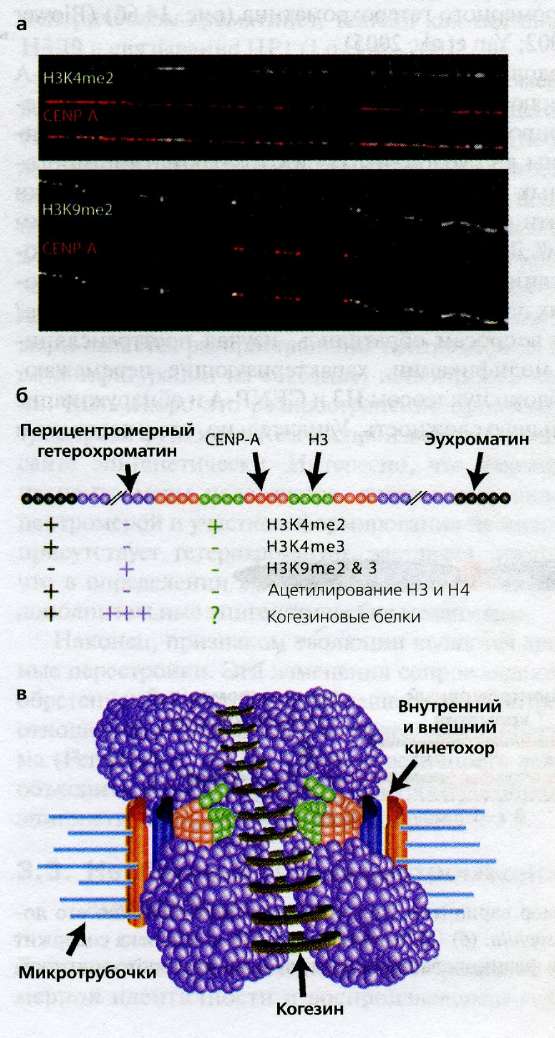
Различные паттерны модификаций гистонов в центромерном хроматине.
(а) Иммунофлуоресцентный анализ с использованием антител, распознающих специфические модификации гистонов на расправленных хромосомных фибриллах, показал, что перемежающиеся блоки нуклеосом, содержащих НЗ, обладают паттерном модификаций, отличающимся от канонических эухроматина и гетерохроматина ( Sullivan and Karpen, 2004 ). Например, несмотря на то, что центромеры у большинства эукариот погружены в крупные блоки перицентрического гетерохроматина, перемежающиеся блоки НЗ содержат модификацию НЗК9mе2, в норме ассоциирующуюся с "открытым" эухроматином (наверху) и лишены гетерохроматинового маркера НЗК9mе2, присутствующей в перицентрических фланкирующих участках (внизу).
(б) Сводка "двумерной" организации центромерного хроматина в интерфазе, основывающаяся на исследованиях растянутых фибрилл хроматина у мух и человека. + и - обозначают присутствие и отсутствие указанных модификаций гистонов (соответственно) в эухроматине, перицентромерном гетерохроматине и перемежающихся блоках нуклеосом НЗ в центромерном хроматине ( Sullivan and Karpen, 2004 ; Lam et al., 2006 ).
(в) Модель трехмерной организации хроматина в центромерном районе митотических хромосом. Предполагается, что ассоциации между сходным образом модифицированными нуклеосомами вносят вклад в формирование отдельных трехмерных структур в центромерном и фланкирующем хроматине. Перемежающиеся CENP-A/CID и по-разному модифицированные НЗ и Н4 могут опосредовать формирование "цилиндрических" трехмерных структур, наблюдаемых в метафазных хромосомах ( Blower et al., 2002 ; Sullivan and Karpen, 2004 ). Хроматин H3K9me2, рекрутирующий такие белки гетерохроматина, как НР1, и такие белки когезии, как RAD21/SCC1, присутствует во внутреннем кинетохорном пространстве между митотическими сестринскими хроматидами и в участках, фланкирующих центромерный хроматин. Это расположение может быть необходимо для "презентации" CENP-A направленной к полюсу поверхности митотической хромосомы и облегчения рекрутирования внешних кинетохорных белков, а также для стимуляции взаимодействия НР1 с самим собой и правильной конденсации/когезии хромосом. Когезины изображены в виде кольцевидных структур в соответствии с последними моделями.