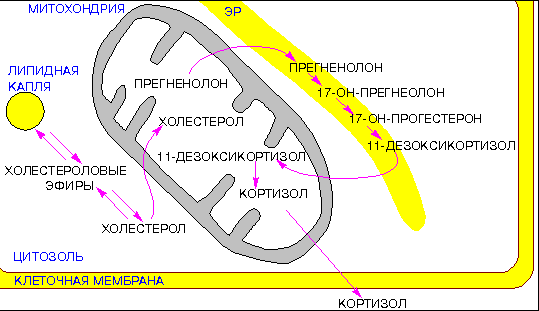
Рис. 4.6(epig). Этапы сборки теломерного гетерохроматина
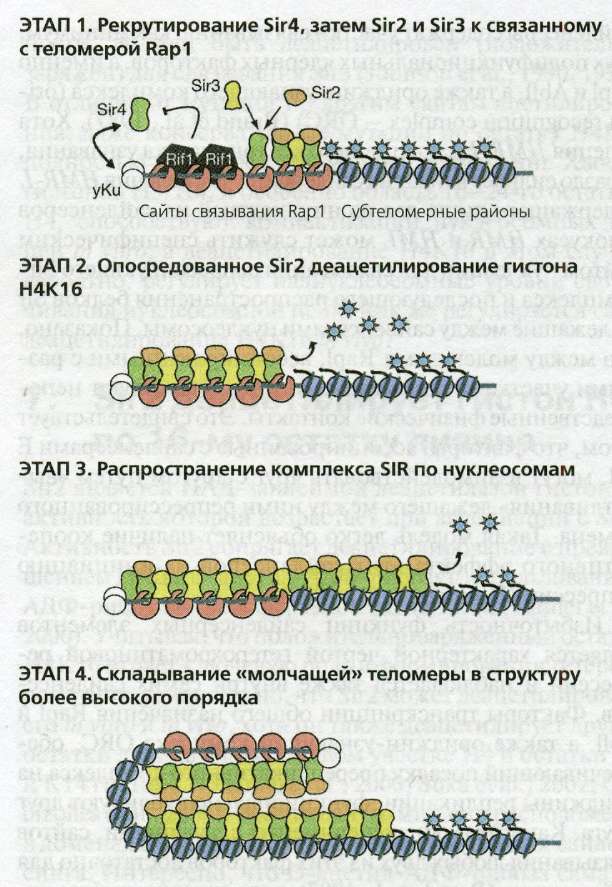
ЭТАП 1. Рекрутирование Sir4, затем Sir2 и Sir3 связанным с теломерами Rap 1. На теломерах Rap 1 и yKu рекрутируют Sir4 даже в отсутствии Sir2 и Sir3. Только Sir4 может быть рекрутирован в отсутствии остальных Sir-белков, причем его связыванию препятствуют Rif1 и Rif2 ( Mishra and Shore 1999 ).
ЭТАП 2. Опосредованное Sir2 деацетилирование гистона Н4К16. Sir4-Sir2 и Sir4-Sir3 активно взаимодействуют, образуя Sir-комплексы по длине TG-повторов. НАД-зависимая гистон-деацетилазная активность Sir2 стимулируется образующимися комплексами и деацетилирует К16 остаток гистона Н4 в близлежащих нуклеосомах.
ЭТАП 3. Распространение SIR-комплекса по нуклеосомам. SIR-комплексы распространяются по нуклеосомам, возможно, с использованием О-ацетил-АДФ-рибозных интермедиатов, образующихся в результате НАД-гидролиза ( Liou et al. 2005 ). Sir3 и Sir4 связываются с деацетилированными хвостами гистона Н4. Хотя деацетилированный N-концевой хвост гистона НЗ также связывает белки Sir3 и Sir4, он не показан.
ЭТАП 4. Свертывание "молчащей" теломеры в структуру более высокого уровня. "Молчащий" хроматин "созревает" в конце М-фазы с образованием недоступной структуры. Этот процесс может включать более высокие уровни свертывания и связывание с ядерной оболочкой.
Ссылки:
- Образование теломерных петель у S. cerevisiae
- Теломерный гетерохроматин
- Sir2 Деацетилирует гистон Н4 по 16-му остатку лизина у S. cerevisiae
- Нарушения непрерывности репрессии теломерными петлями у S. cerevisia
- Отдельные этапы сборки гетерохроматина: общие сведения
- Рис. 4.8(epig). Организация природных теломер и характер их сайленсинга