Эпигенетический контроль инициации репликации ДНК
Инициация репликации ДНК контролируется эпигенетическими механизмами.
Надежное копирование ДНК осуществляется ДНК-полимеразами, действующими от 5'- к З'-концу, и начинается в специфических сайтах, называемых "ориджинами" (точками начала репликации ДНК). В "ориджинах" образуется "пузырь", состоящий из двух репликационных "вилок", и репликация идет в двух направлениях, пока не встречаются "вилки", генерируемые следующими "ориджинами". Домены, реплицируемые с одного "ориджина", обычно очень большие, длиной в сотни тысяч пар оснований ( Aladjem and Fanning, 2004 ). "Ориджины" у дрожжей Saccharomyces cerevisiae (известные также как ARSs - autonomously replicating sequences, автономно реплицирующиеся последовательности) функционируют эктопически, будучи клонированы в плазмиды, и обычно после интеграции в другие хромосомные сайты, показывая тем самым, что инициация репликации регулируется специфическими последовательностями нуклеотидов ДНК. ARSs имеют величину приблизительно 100-150 п.н. и содержат одну или несколько копий важной последовательности длиной приблизительно 11 п.н. и обогащенной AT плюс другие консервативные элементы ( Weinreich et al., 2004 ). Для инициации необходим белковый комплекс, известный как комплекс распознавания "ориджинов" ( ORC , origin recognition complex), который отвечает за рекрутирование предрепликационного комплекса ( PRC , prereplication complex), включающего белки поддержания минихромосом ( MCM , minichromosome maintenance proteins) ( Prasanth et al., 2004 ). У многоклеточных "ориджины" репликации можно идентифицировать in situ, но обычно после клонирования и реинтродукции они не активны, что позволяет предполагать, что у этих организмов инициация регулируется эпигенетически, а не строгой зависимостью от последовательности ДНК ( Aladjem and Fanning, 2004 ). Хотя у многоклеточных белки ORC и МСМ консервативны, факторы и механизмы, ответственные за регуляцию активности "ориджинов" у этих организмов, остаются загадкой. Кроме того, неясно, влияет ли структура хроматина на работу репликационных "вилок".
Ключ к тому, каким образом "ориджины" многоклеточных могли бы регулироваться эпигенетически, дают детальные исследования на S. cerevisiae. Хотя нуклеотидные последовательности ДНК в "ориджинах" необходимы и, по большей части, достаточны для инициации репликации у этого организма, имеются четкие данные о влиянии структуры хроматина на активность "ориджинов" ( Weinreich et al., 2004 ). Анализ с использованием микрочипов (microarray analysis) показал, что не все из 332 сайтов двунаправленной репликации или из 429 сайтов связывания белков ORC и МСМ активны в каждом клеточном цикле. Хромосомный контекст и структура хроматина влияют на способность предполагаемого "ориджина" быть активным в инициации репликации. Например, ARS, локализованная в "молчащих" локусах типа спаривания, не может инициировать репликацию, если не будет перемещена в другие места хромосомы. Другой пример эпигенетической регуляции "ориджинов" относится к приблизительно 100-200 генам, кодирующим рибосомную РНК (rRNA) , которые расположены в виде тандемных повторов ( Pasero et al., 2002 ). Каждая единица rDNA длиной 9.1 т.п.н. содержит ARS, которая инициирует репликацию, будучи вставлена в плазмиду или в другое место генома. Однако в каждой S-фазе активны лишь около 20% "ориджинов" rDNA.
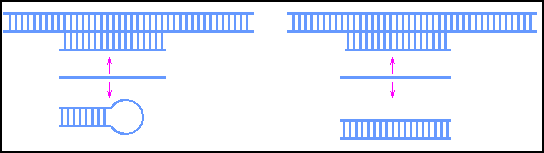
"Ориджины" могут также регулироваться таким образом, что будут работать [to fire] на разных стадиях S-фазы ( Weinreich et al., 2004 ). Например, "ориджины" возле теломер обычно активны в поздней S-фазе (см. " Эпигенетический контроль структуры и функции теломер "), но вставка тех же самых последовательностей в кольцевые плазмиды приводит к репликации в ранней S-фазе ( рис. 14.3 ) ( Ferguson and Fangman, 1992 ). Поздняя репликация может рекапитулировать в результате линеаризации такой плазмиды и добавления теломер, что помещает ARSs вблизи концов.
Эти результаты показывают, что как активность "ориджинов", так и время начала работы "ориджина" в ходе S-фазы регулируются эпигенетически. Дальнейшие доказательства вытекают из наблюдений, согласно которым "расписание" S-фазы, сайленсинг "ориджинов" в rDNA, теломеры и локусы типа спаривания регулируются многими из модификаций гистонов и белками, отвечающими за эпигенетический сайленсинг генов, в том числе белками SIR (глава " ЭПИГЕНЕТИКА ДРОЖЖЕЙ S. CEREVISIAE ").