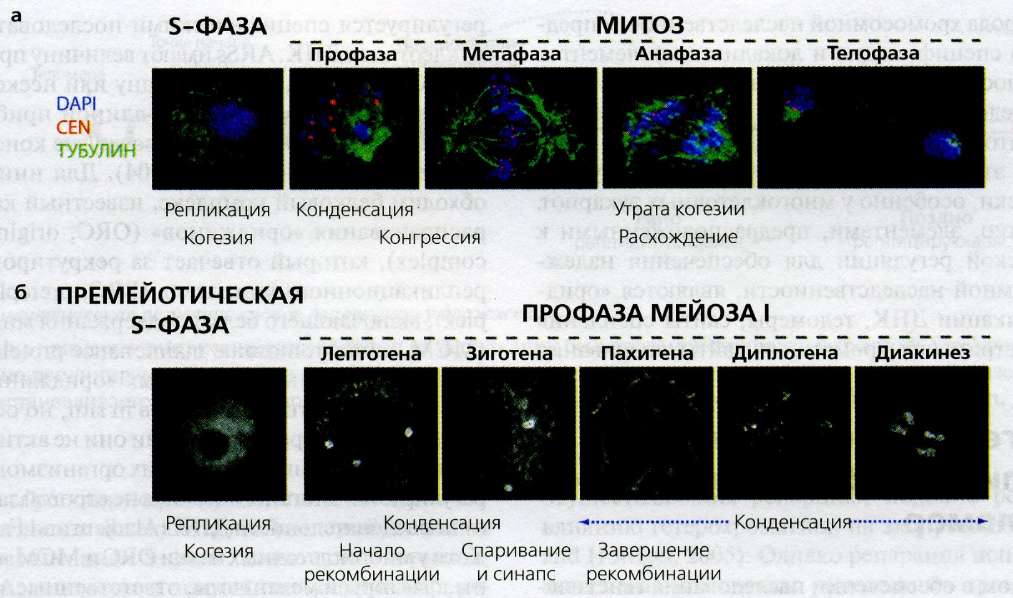
Эксцизионная репарация нуклеотидов (ЭРН) E. coli
У E. coli эксцинуклеаза формируется, в результате димеризации двух молекул белка UvrА в присутствии АТР и связывания с одной молекулой UvrB. Гетеротример UvrA2B связывается с ДНК и с помощью 5'-3'-ДНК-геликазной активности перемещается вдоль ДНК в поисках повреждения. Такова предполагаемая картина узнавания эксцинуклеазой UvrAВС неспецифического повреждения в ДНК [ Hoeijmakers J., 1993 ]. Результатом узнавания повреждения этим комплексом явится внедрение субъединицы UvrB в ДНК, сопровождающееся конформационными изменениями ДНК в сайте внедрения ( изломы , локальная денатурация), диссоциация обеих субъединиц UvrА из комплекса и последующее связывание с ним молекул UvrD и UvrC. Гетеродимер UvrBC из состава нового комплекса делает два разрыва в поврежденной нити ДНК на расстоянии 8 н. с 5'-конца от повреждения и 5 н. с 3'-конца, катализируемых субъединицами UvrC и UvrB соответственно, что приводит к появлению 12-13-мерного олигонуклеотида, вытесняемого из комплекса с помощью геликазы UvrD. Образующаяся при этом брешь заполняется ДНК-полимеразой I, синтезирующей ДНК по неповрежденной матрице, и сшивается ДНК-лигазой. Описанный процесс зависит от АТР. Действительно, АТР стимулирует димеризацию молекул UvrA, гидролиз АТР необходим для образования комплекса UvrA2B и проявления его геликазной активности, АТР необходим для формирования комплекса UvrB-ДНК и, наконец, для описанной выше бимодальной инцизии ДНК [ Friedberg E.C., Walker G.C., ea., 1991 ]. У E. coli транскрипционно-зависимую ветвь ЭРН (ТЭРН) осуществляет продукт гена mfd (mutation frequency decline). Этот ген был открыт за 35 лет до того, как его продукт признали фактором TRCF. Последний способствует диссоциации РНК-полимеразы от дефектной транскрибируемой нити ДНК, и связывается с субъединицей UvrA эксцинуклеазного комплекса. Иными словами, именно белок Mfd отыскивает повреждения на транскрибируемой нити и направляет ТЭРН на эту мишень. Согласно недавним наблюдениям, два главных белка системы ДКНО - MutL и MutS - необходимы для ТЭРН [Melon I., Champl G.N., 1996]. Причина такой взаимосвязи систем ДКНО и ТЭРН или заимствования белков MutL и MutS для выполнения сходных функций пока остается загадочной, хотя сам факт несомненен и нашел подтверждение также и для белков Msh2, Mlh1, Pms1 и Msh3 у S. cerevisiae [ Sweder K.S., ea, 1996 ].