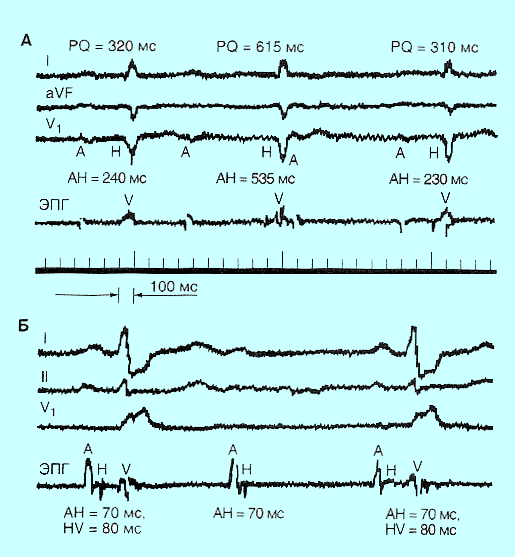
Ген c-fos: регуляция, пути передачи сигнала, общее представление
Схема путей активации гена c-fos представлена на рис.3 . Говоря в общем, мы наблюдаем взаимозависимое действие разных путей передачи сигнала. С одной стороны, МАР-киназный путь приводит к фосфорилированию факторов тройного комплекса и активации транскрипции c-fos через промоторный элемент SRE .
С другой стороны, ERK -зависимое фосфорилирование c-Jun по С-концевому домену приводит к понижению его ДНК-связывающей активности и, как следствие, к ингибированию TRE -опосредуемой активации транскрипции.
В то же время ТРА , стимулирующий РКС , вызывает, вероятно, активацию фосфатаз , дефосфорилирование c-Jun и активацию АР-1 комплекса .
При этом следует иметь ввиду, что прочное связывание транскрипционных факторов ( SRF/TCF и АР-1 ) с двумя регуляторными элементами SRE и TRE является взаимоисключающим из-за стерических ограничений [ Lucibello F.C. ea, 1991 ]. Тем не менее эффекты ТРА распространяются помимо ТRE также и на SRE ( рис.1 ) [ Sasson-Corsi P. ea,1989 ].
Скорее всего, тип активируемого цисэлемента определяется конкретными внутриклеточными условиями, но в любом случае приводит к активации транскрипции.
С другой стороны, часто индукция гена c-fos сопровождается стимуляцией ERK , а не JNK [ Minden A. ea, 1994 , Su B. ea, 1994 ]. Возможно, что элемент SRE является главной мишенью для путей передачи сигнала на промотор гена c-fos [ Lucibello F.C. ea, 1991 ].
Возникает вопрос о необходимости такой регуляции генной экспрессии, когда стимуляция транскрипции через один элемент подразумевает ингибирование действия другого активирующего элемента.
Кажется противоречивым, что инактивация МАРККК ( c-Raf ) блокирует трансформацию клеток форболовыми эфирами [ Hunter T., 1991 ], поскольку блокируемое при этом стимулирование транскрипции через SRE, казалось бы, должно восполняться активацией TRE. Ответ следует из того факта, что SRE гораздо более эффективен при транскрипционной активации, чем TRE.
Более того, TRE может служить и в качестве ингибитора SRE-активности, поскольку устранение АР-1 сайта из промоторной области повышает базальную активность транскрипции c-fos [ Lucibello F.C. ea, 1991 ]. Вероятно поэтому уровень экспрессии гена c-fos при индукции его с помощью ТРА с параллельным ингибированием МАРККК недостаточен для трансформации клеток.
Таким образом, на данном уровне осуществляется регуляция активности гена c-fos в зависимости от характера воздействия.
Остается вопрос, почему ТРА , активирующий РКС , может индуцировать транскрипцию c-fos, используя два промоторных элемента ( SRE и TRE ), в то время как другие факторы, также активирующие РКС, используют только один промоторный элемент SRE. Вероятно, разные факторы активируют определеннные изоформы РКС , тогда как активирующее действие ТРА распространяется на большинство членов РКС-семейства. Кроме того, для передачи сигнала на элемент SRE необходимо сложное взаимодействие многих мембранных и цитоплазматических компонентов, что в конечном счете приводит к активации строго определенной группы транскрипционных факторов, тогда как непосредственное действие форболовых эфиров на РКС ограничивает возможности тонкой регуляции, что проявляется в более широком спектре активируемых факторов.
Благодаря регулированию стабильности м-РНК активность гена c-fos может тонко регулироваться несколькими РКС-зависимыми путями. Набор изоформ, входящих в семейства факторов, составляющих путь передачи сигнала (GTP- связывающих белков, фосфолипаз, РКС, МАР-киназ), делает возможным такую многовариантную регулировку гена c-fos и, по всей вероятности, многих других генов. Это создает основу для точного включения гена c-fos в контекст внутриклеточных событий. Однако многие подобные события регулируются с участием других сигналпередающих систем. Ген c-fos также находится под контролем и этих систем, в частности cAMP-зависимых путей передачи сигнала.