Сенсорное преобразование (трансдукция) в рецепторах
Сенсорные системы организованы таким образом, что они реагируют на окружающую организм среду. Событие в среде, возбуждающее сенсорный рецептор, - это стимул. Информацию о стимуле рецепторы направляют в ЦНС. Эффект, вызываемый им в организме, называется ответом.
Ответы можно рассматривать на разных функциональных уровнях нервной системы: это рецепторные потенциалы в сенсорных клетках; последовательности (серии) потенциалов действия (импульсов) в аксонах сенсорных путей; синаптические явления в сенсорных нейронных сетях; двигательная активность, вызываемая сенсорными стимулами; наконец, поведенческие акты. Процесс, обеспечивающий адекватный ответ сенсорного рецептора на стимул, называется сенсорным преобразованием (трансдукцией).
События окружающей среды, запускающие сенсорное преобразование, всегда представляют собой форму энергии - механическую, тепловую, химическую. Каждый тип сенсорного рецептора специализируется на преобразовании одной формы энергии. Некоторые организмы способны преобразовывать такие формы энергии, которые недоступны для восприятия другими организмами. Например, человек не ощущает электрические и магнитные поля, тогда как у многих рыб есть электрорецепторы, а рыбы и птицы ориентируются во время своих миграций по магнитному полю Земли.
Во всех сенсорных рецепторах преобразование начинается с вызванного стимулом изменения свойств клеточной мембраны. На рис. 33.2 показано, каким образом под действием разных стимулов изменяются свойства мембраны сенсорных нейронов, специализированных для преобразования этого вида стимулов. Например, ответ хеморецептора ( рис. 33.2 , а) возникает при взаимодействии молекулы стимулирующего химического вещества с рецепторными молекулами плазматической мембраны сенсорного нейрона. (Обратите внимание на различие понятий: сенсорный рецептор состоит из одной или нескольких клеток, тогда как рецепторная молекула - это интегральный белок клеточной мембраны). В результате реакции между стимулирующим химическим веществом и рецепторными молекулами мембраны открываются ионные каналы, через которые в клетку входит поток ионов, обеспечивающий деполяризацию сенсорного рецептора. На рис. 33.2 ,б показано открывание ионного канала механорецептора в ответ на механическую силу, направленную вдоль мембраны (растяжение). На рис. 33.2 , в представлен ионный канал фоторецептора - клетки, реагирующей на свет. В темноте канал находится в открытом состоянии. Когда особым пигментом мембранного диска наружного сегмента рецептора поглощается фотон света, он закрывается. Таким образом, входящий ток возникает в темноте и соответственно называется темновым током ; при освещении глаза он прекращается. Исчезновение тока сопровождается гиперполяризацией фоторецептора.
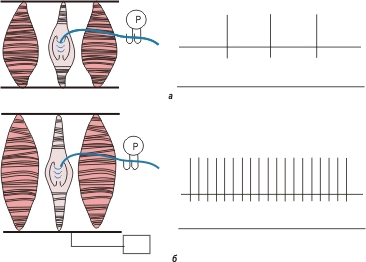
Сенсорная трансдукция приводит, как правило, к генерированию рецепторного потенциала в периферическом окончании сенсорного афферентного нейрона ( рис. 33.3 ). Обычно это деполяризация, обусловленная входящим ионным током; она сдвигает мембранный потенциал сенсорного рецептора к пороговому уровню запуска потенциала действия. Например, механический стимул ( рис. 33.3 ) деформирует механорецепторы, вызывая в окончании аксона входящий ток ионов, которые распространяются в продольном направлении внутри аксона и выходят наружу из более проксимальных его участков. Выходящий ионный ток деполяризует мембрану, т.е. возникает рецепторный потенциал; его амплитуда может превысить пороговый уровень запуска потенциала действия. В нашем примере потенциал действия генерируется в триггерной зоне первого перехвата Ранвье афферентного волокна.
В некоторых сенсорных образованиях рецепторный потенциал, генерируемый в результате сенсорной транедукции, представляет собой не деполяризацию, а гиперполяризацию. Например, гиперполяризующий рецепторный потенциал характерен для фоторецепторов.
Существуют сенсорные образования, в которых окончание первичного афферентного волокна само по себе не является рецептором, а контактирует с особой периферически расположенной сенсорной клеткой. Например, первичные афференты улитки внутреннего уха образуют окончания на волосковых клетках . В таких сенсорных образованиях трансдукция происходит более сложно. Звук вызывает в волосковых клетках улитки колебательный рецепторный потенциал. Во время каждого его колебания (осцилляции) мембрана волосковой клетки деполяризована. В результате этого из волосковой клетки высвобождается возбуждающий нейромедиатор. Он взаимодействует с мембраной окончания первичного афферента, вызывая, в свою очередь, деполяризующий генераторный потенциал, который сдвигает мембранный потенциал первичного афферента по направлению к пороговому уровню запуска нервного импульса.