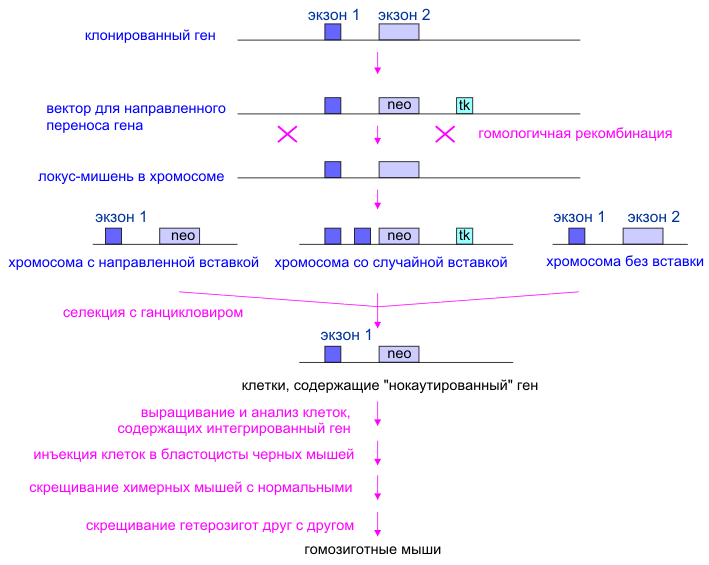
Рис. 5. rib Динамическая модель работы рибосомы в элонгационном цикле
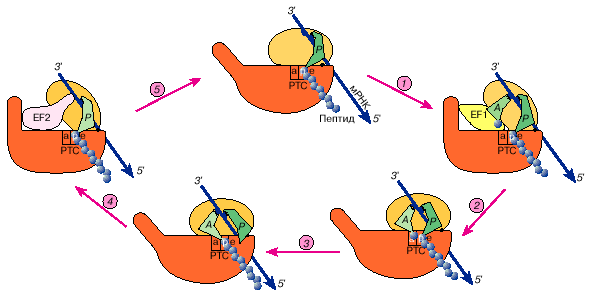
На данной схеме "головки" обеих рибосомных субчастиц обращены к зрителю. Модель постулирует, что две рибосомные субчастицы подвижно соединены друг с другом и способны к некоторому раздвиганию (размыканию) и сдвиганию (смыканию). Размыкание открывает функциональные центры на контактирующих поверхностях субчастиц, такие, как А-участок для приема аминоацил-тРНК, и способствует перемещению лигандов, например в ходе транслокации. Смыкание субчастиц запирает лиганды внутри рибосомы и приводит в тесный контакт субстраты для реакции транспептидации. Предполагается, что размыкание индуцируется факторами элонгации с ГТФ, а гидролиз ГТФ и уход факторов с рибосомы позволяют ей снова сомкнуться. На рисунке представлено рассмотрение элонгационного цикла с точки зрения модели смыкания-размыкания:
1- связывание аминоацил-тРНК с рибосомой требует размыкания, и фактор элонгации EF1 с ГТФ призван "открыть" рибосому;
2 - после расщепления ГТФ фактор элонгации EF1 покидает рибосому, а аминоацильный конец аминоацил-тРНК взаимодействует с пептидилтрансферазным центром большой субчастицы и тем самым способствует смыканию субчастиц и запиранию рибосомы;
3 - реакция транспептидации происходит в "закрытой" рибосоме между тесно сближенными группами двух субстратов - пептидил-тРНК и аминоацил-тРНК;
4 - размыкание претранслокационной рибосомы промотируется фактором элонгации EF2 с ГТФ, что приводит к выходу деацилированной тРНК и смещению остатка тРНК молекулы пептидил-тРНК вместе с мРНК;
5 - гидролиз ГТФ и как следствие уход фактора элонгации EF2 снова дает возможность рибосоме сомкнуться. Таким образом, транслирующая рибосома в ходе элонгационного цикла осциллирует между сомкнутым и разомкнутым состояниями