Биофизика волосковых клеток улитки: общие сведения
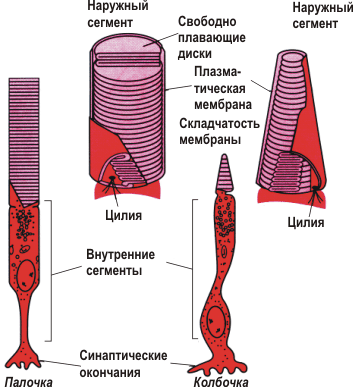
Мы отмечали, что волосковые клетки улитки подразделяются на два типа: наружные и внутренние ( рис. 8.13 ). Они различаются в разных частях улитки и по размерам. У основания улитки морской свинки наружные волосковые клетки имеют длину около 20 мкм и короткие неэластичные стереоцилии , тогда как на вершине - 80 мкм с более длинными и гибкими стереоцилиями. Эти различия в длине сказываются не только на их механических, но и электрических свойствах. Показано, что на плазматических мембранах волосковых клеток наблюдается спонтанная активность. Частота этих осцилляций варьирует в различных частях улитки. Вблизи от круглого окна, у основания улитки, в маленьких неэластичных наружных волосковых клетках короткие неэластичные наружные волосковые клетки демонстрируют быстрые спонтанные осцилляции, дальше в направлении геликотремы более длинные клетки - более медленные. Существует зависимость между размером и частотой спонтанной электрической активности клетки . Поскольку размеры наружных волосковых клеток закономерно изменяются от основания к верхушке улитки, спонтанные осцилляции соответствуют звуковой частоте, на которую настроена подлежащая базиллярная мембрана . Таким образом, когда поступающий слуховой сигнал индуцирует механическую вибрацию основной мембраны, возникающие вследствие открывания и закрывания ионных каналов стереоцилий электротонические токи, усиливают предсуществующую активность. Это явление известно как электрический резонанс. Волосковая клетка работает как усилитель, повышая чувствительность уха на два порядка. Различия электрического резонанса от основания к верхушке улитки подразумевают, что популяция ионных каналов в волосковых клетках также закономерным образом меняются от круглого окна к геликотреме. Такое предположение подтверждается установлением того факта, что количество каналов Са2+ и каналов Са2+-зависимых К+ (КСа) уменьшается со снижением характеристической частоты. Исследование улитки цыпленка показало, что каждая из приблизительно 10 000 волосковых клеток обладает уникальной популяцией ионных каналов. Причем клетки различаются не только количеством каналов, но и биофизическими характеристиками последних. Прорыв был сделан в молекулярной генетике этой системы. Показано, что различные К+-каналы возникают в результате альтернативного сплайсинга в семи различных сайтах первичных транскриптов. Электрический резонанс - это не только механизм, благодаря которому наружные волосковые клетки увеличивают точность настройки. Показано, что электрический ответ связан с механическим. Деполяризация ведет к укорочению волосковой клетки, гиперполяризация имеет противоположный эффект, вызывая удлиннение клетки. В точке максимальной чувствительности такая реакция может достигать 30 нм/мВ. Механизм трансдукции изменений мембранного потенциала наружных волосковых клеток в изменения длины до сих пор неизвестен. Однако очевидно, что и это повышает специфичность клетки к частоте звука. Чем выше электрический резонанс, тем больше механическое движение, что ведет к увеличению натяжения между стереоцилиями и текториальной мембраной. В свою очередь, это приводит к повышению вероятности открытого состояния механочувствительных каналов стереоцилий. Полагают, что система может работать и в противоположном направлении. Спонтанные или индуцированные афферентами движения волосковых клеток могут вызывать движения основной мембраны и изменения давления в средней лестнице и далее, которые компенсируются движениями барабанной перепонки. Это - т.н. спонтанная отоакустическая эмиссия , которая может быть обнаружена ухом человека и других млекопитающих. Связаны ли эти движения с "звоном в ушах", до сих пор остается предметом противоречий. Литература (см. список на стр. 109-110 книги)
Теперь обратим внимание на основную тему данного предмета. Мы видели, что базиллярная мембрана колеблется в ответ на поступающий в ухо звук, тогда как текториальная мембрана остается сравнительно стационарной. Стереоцилии волосковых клеток подвергаются механической деформации, причем их реснички погружены в богатую К+ эндолимфу . Возникающая деполяризация может быть обнаружена с помощью микроэлектродных отведений. Они точно воспроизводят частоту поступающего звука. Это т.н. микрофонные потенциалы . Микрофонные деполяризации (рецепторные потенциалы) ведут к выделению медиаторных веществ на дендритные окончания афферентных волокон кохлеарного нерва.
Таким образом, мы видим, что в самой основе потрясающе сложного внутреннего уха млекопитающих лежат волосковые клетки; конечно, модифицированные, но в целом те же самые, что мы впервые встретили в каналах органа боковой линии наших водных предшественников. Мы еще увидим, что примерно тоже самое можно сказать и о других органах чувств. Молекулярные механизмы, развившиеся в эволюционной истории очень рано, сохраняются, но со временем оказываются встроенными в невероятно сложные и хитроумные органы. Одним из эволюционных императивов, который двигал развитием улитки млекопитающих, была необходимость различать различные частоты звука. Мы видели, что эта способность в небольшой степени присутствует у рыб, земноводных и пресмыкающихся; у птиц и млекопитающих она претерпевает огромное развитие. Выше мы упоминали, что частотный диапазон человеческого уха лежит между 20 Гц и 20 кГц (с некоторым снижением верхнего предела с возрастом). Мы также отмечали, что в пределах диапазона слышимости человек и другие млекопитающие обладают чрезвычайно высокой способностью к различению частот. А потому следующий вопрос - как она достигается? Может показаться, что у этой проблемы - простое решение. Почему бы кохлеарному нерву не быть фазово-синхронным с приходящей звуковой волной давления? Иными словами, почему бы о тональной частоте 20 Гц не сигнализировать нервными импульсами частотой 20 Гц, а о тональной частоте 15 или 20 кГц - импульсами частотой 15 и 20 кГц, соответственно? В таком простом решении есть две очевидные трудности. Во-первых, как мы отмечали в гл МЕМБРАНЫЕ ПОТЕНЦИАЛЫ , частота импульсов в сенсорных нервах обычно сигнализирует об интенсивности стимула. Нервная система могла бы, конечно, обойти это затруднение, однако, вторая трудность- более непреодолима. Биофизика нервных волокон такова, что за каждым импульсом следует рефрактерный период длительностью около 2 мс. Из этого вытекает (как мы видели в гл. МЕМБРАНЫЕ ПОТЕНЦИАЛЫ ), что одиночное волокно не способно проводить более 500 импульсов в секунду. Т.е., для частот выше 500 Гц нужны какие-то иные средства частотной дискриминации. Здесь могут работать два основных механизма. Во-первых, есть данные (см. гл. АНАЛИЗ ВЕСТИБУЛЯРНОЙ И ЗВУКОВОЙ ИНФОРМАЦИИ В МОЗГЕ ), что кохлеарные волокна могут быть фазо-синхронными звуковым частотам выше 500 Гц, но, не реагируя на каждый частотный импульс. То есть, предполагается, что в нижней части частотного спектра (ниже 5 кГц) группа волокон кохлеарного нерва объединяется для достижения частоты импульсов, совпадающей с тональной частотой, в каком-то слуховом центре мозга. По понятным причинам, такая идея именуется теорией залпа. Второй, значительно более важный механизм основывается на наблюдении, что ширина основной мембраны увеличивается от круглого окна к геликотреме (или в случае птиц - к макуле улитки). Ширина основной мембраны человека, например, увеличивается с 100 до 500 мкм на расстоянии в 33 мм ( рис. 8.17 ). Герман фон Гельмгольц еще в ХIX веке предположил, что основную мембрану можно уподобить ряду настроенных камертонов (резонаторов). Тоны высокой частоты вызывают максимальные возмущения в области круглого окна, а низкой - у геликотремы. Точные исследования фон Бекеши (von Bekesy) и других в основном подтвердили гипотезу Гельмгольца. Обнаружено, что волны сложной формы двигаются вдоль всей основной мембраны, но место, где они достигают максимальной амплитуды, как и предположил Гельмгольц, связано с их частотой. Догадка Гельмгольца по очевидным причинам известна как теория места частотной дискриминации . Чтобы различить частоты, мозгу достаточно лишь "посмотреть" из какого места основной мембраны происходят волокна, в которых активность максимальна.