Эволюция растений
Появление наземных, или высших, растений ознаменовало начало новой эры в жизни нашей планеты. Освоение растениями суши сопровождалось появлением новых, наземных, форм животных; сопряженная эволюция растений и животных привела к колоссальному разнообразию жизни на земле, изменила ее облик. Первые достоверные наземные растения, известные только по спорам , датируются началом силурийского периода . Из верхнесилурийских и нижнедевонских отложений по сохранившимся макроостаткам или по отпечаткам органов описаны наземные растения. Эти первые известные нам высшие растения объединены в группу риниофитов . Несмотря на анатомическую и морфологическую простоту строения, это были уже типичные наземные растения. Об этом свидетельствует наличие кутинизированной эпидермы с устьицами, развитой водопроводящей системы, состоящей из трахеид , и наличие многоклеточных спорангиев с кутинизированными спорами. Следовательно, можно предположить, что процесс освоения суши растениями начался значительно раньше - в кембрии или в ордовике .
Предпосылок для появления наземных растений было, по-видимому, несколько. Во-первых, независимый ход эволюции растительного мира подготовил появление новых, более совершенных форм. Во-вторых, за счет фотосинтеза морских водорослей в атмосфере земли произошло увеличение количества кислорода; к началу силурийского периода оно достигло такой концентрации, при которой оказалась возможной жизнь на суше. В-третьих, в начале палеозойской эры на обширных территориях земли происходили крупнейшие горообразовательные процессы, в результате которых возникли Скандинавские горы, горы Тянь-Шань, Саяны. Это вызвало обмеление многих морей и постепенное появление суши на месте бывших мелких водоемов. Если раньше водоросли , населявшие литоральную зону, только в отдельные кратковременные периоды жизни оказывались вне воды, то по мере обмеления морей они переходили к более длительному пребыванию на суше. Это, очевидно, сопровождалось массовой гибелью водорослей; выживали лишь те немногие растения, которые смогли противостоять новым условиям жизни.
В ходе длительного эволюционного процесса возникали новые виды, постепенно формировавшие типичные наземные растения.
К сожалению, промежуточные формы палеонтологическая летопись не сохранила. Новая воздушно-наземная среда обитания оказалась чрезвычайно противоречивой, принципиально отличной от первоначальной водной. Прежде всего она характеризовалась повышенной солнечной радиацией, дефицитом влаги и сложными контрастами двухфазной воздушно-наземной среды. Вполне вероятно допустить, что у некоторых переходных форм в процессе обмена веществ мог вырабатываться кутин , который откладывался, на поверхности растений. Это было первым этапом формирования эпидермы . Чрезмерное выделение кутина неминуемо приводило к гибели растений, так как сплошная пленка кутина препятствовала газообмену. Только те растения, у которых выделялось умеренное количество кутина, смогли сформировать сложную специализированную ткань - эпидерму с устьицами, способную и защищать растение от высыхания, и осуществлять газообмен. Таким образом, важнейшей тканью наземных растений, без которой невозможно освоение суши, следует считать эпидерму. Однако возникновение эпидермы лишало наземные растения возможности поглощать воду всей поверхностью, как это происходит у водорослей.
У самых первых наземных растений, имевших еще небольшие размеры, поглощение воды осуществлялось с помощью ризоидов - одноклеточных или многоклеточных однорядных нитей. Однако по мере увеличения размеров тела происходил процесс формирования сложных специализированных органов - корней с корневыми волосками . По-видимому, образование корней, начавшееся с верхнедевонского периода , в разных систематических группах растений происходило разными путями. Активное поглощение воды ризоидами и корнями стимулировало возникновение и совершенствование водопроводящей ткани - ксилемы . У нижнедевонских растений ксилема состояла только из кольчатых и спиральных трахеид . Начиная с верхнего девона прослеживается тенденция к "оживлению" ксилемы за счет развития древесной паренхимы , которая способствовала более активному проведению воды.
Выход растений на сушу сопровождался улучшением их освещения, что активизировало процесс фотосинтеза . Это приводило к увеличению количества ассимилятов, а следовательно, к увеличению объема растений, что вызвало необходимость их морфологического расчленения.
Существует несколько взглядов на исходную форму первых наземных растений. Одни авторы считают первичной формой пластинчатую - талломную , другие - напротив, радиальную. Есть третья точка зрения, согласно которой первые наземные растения выводятся из гетеротрихальных зеленых водорослей типа хетофоровых . Их стелющиеся части тела дали начало талломным формам, а восходящие - радиальным, т.е. талломные и радиальные структуры возникали одновременно и развивались параллельными путями. Пластинчатые талломы оказались малоперспективными в биологическом отношении, так как они очень быстро опутали бы всю поверхность земли тонким слоем, вызвав обостренную конкуренцию за свет. Восходящие структуры, напротив, получили дальнейшее развитие и сформировали радиальные ветвящиеся осевые органы. Вертикальное положение тела растения оказалось возможным только при условии возникновения механических тканей . (Лигнификация клеток явилась, по- видимому, следствием избытка углеводов в ходе усиленного фотосинтеза.)
Независимо от формы тела во всех группах наземных растений очень рано (с середины девона ) начала проявляться тенденция к образованию плоских боковых фотосинтезирующих органов - листьев . Подобно корням листья возникали разными путями, т.е. в разных систематических группах они имеют разное происхождение. (Своеобразие происхождения листьев нашло отражение в терминологии; так, все листья мохообразных иногда называют филлидами , листья плауновидных - микрофиллами, или филлоидами , листья папоротникообразных - макрофиллами, или вайями . Однако эти термины не всегда раскрывают специфику листьев разных групп растений.)
Большая поверхность листьев в сочетании с наиболее совершенной, зернистой формой хлоропластов способствовала увеличению эффективности процессов фотосинтеза , т.е. накоплению органического вещества. Быстрое и равномерное распределение пластических веществ по всему телу растения оказалось возможным только при наличии совершенной проводящей ткани - флоэмы , которая обнаруживается уже у нижнедевонских риниофитов .
Таким образом, у высших растений произошло формирование важнейших тканей - эпидермы , ксилемы , флоэмы , механической ткани и важнейших органов - листостебельного побега и корня . По-латыни побег - кормос (cormos) , поэтому многие авторы называют высшие растения кормофитами. Однако А.Л.Тахтаджян справедливо отрицает универсальность этого названия, так как среди высших растений есть талломные мохообразные , а первые наземные растения - риниофиты еще не имели сформированных побегов.
Появившись в нижнем палеозое , высшие споровые растения стали быстро распространяться по поверхности земли, осваивая новые территории. Этому в значительной степени способствовало отсутствие в тот период конкуренции. Палеозойская эра характеризовалась интенсивными видообразовательными процессами, что привело к расцвету папоротникообразных , членистых , плауновидных . В мезозойскую эру господство в растительном мире переходит к голосеменным растениям , а в сменившую ее кайнозойскую эру расцвета достигают покрытосеменные растения .
К сожалению общее дерево растений пока не представлено в данной базе данных.
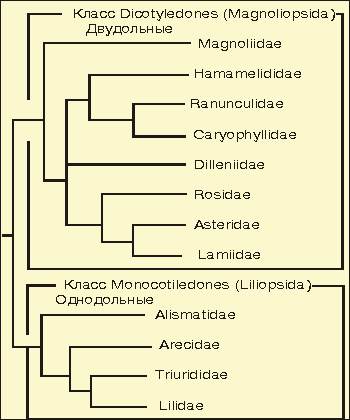
Общее дерево высших растений ( Тахтаджан, 1987 ):