Генетическая предрасположенность или генетический предрак
Опухоль, являющаяся клоном клеток со стабильными патологическими свойствами, является, скорее всего, генетически измененным клоном, результатом одной или ненескольких мутаций. Наследственные формы рака или гемобластозов соответствуют этой точке зрения. Самым ярким примером наследственного рака является ретинобластома - злокачественная опухоль сетчатки глаза [ Weinberg, 2006e ]. Эта опухоль вызывается рецессивной мутацией, передающейся от родителей. Наличия одной функциональной копии гена Rb достаточно для поддержания нормального клеточного фенотипа, однако происходящая спорадически в клетках сетчатки глаза мутация, инактивирующая второй аллель Rb, ведет к возникновению в раннем детстве ретинобластомы; при этом практически все случаи наследственной ретинобластомы характеризуются последовательным поражением обоих глаз [ Knudson, 1971 ]. Следовательно, исходная мутация определяет предрак , реализующийся в большой популяции клеток, где возникает вторая мутация.
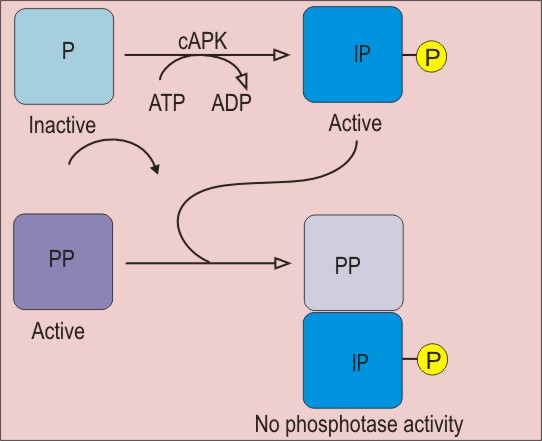
Природа гена Rb в настоящее время установлена. Белковый продукт гена Rb контролирует продвижение клетки по циклу деления. Регуляция функции pRb (белка, кодируемого геном Rb) осуществляется путем его фосфорилирования-дефосфорилирования [ Weinberg, 2006e ]. В неделящихся клетках (G0) pRb дефосфорилирован. В G1 он постепенно фосфорилируется и в гиперфосфорилированном состоянии пересекает "точку рестрикции", отделяющую G1 от S-фазы - фазы синтеза ДНК. Затем pRb дефосфорилируется - до начала нового митотического цикла. Активность фосфорилирования определяется циклином D , взаимодействующим с митогенными сигналами. Мутации pRb делают этот белок независимым от митогенных сигналов, создающим непрерывное (и потому нерегулируемое) прохождение клеток сетчатки по циклу, что лежит в основе возникновения ретинобластомы .
Четкий пример роли генетических изменений показан при исследовании рака молочной железы [ Calderon-Margalit, 2004 ]. Частота возникновения этого рака отчасти контролируется генами BRCA1 и BRCA2 , частота мутаций которых коррелирует с частотой появления этого рака. Природа связи BRCA1 и BRCA2 с возникновением рака молочных желез не установлена, но роль генетических факторов в возникновении этой опухоли в некоторых популяциях весьма вероятна.
Другой яркий пример роли генетических изменений в предраке - динамика гена APC в возникновении рака толстой кишки : выпадение функции гена APC ведет к резкому увеличению риска возникновения аденоматозного полипоза , на фоне которого возможно развитие клона колоректального рака [ Vogelstein, 1988 ]. Однако мутации гена APC самой по себе недостаточно для возникновения злокачественной опухоли. Аденоматозный полипоз создает популяцию высокого риска развития моноклонального рака толстого кишечника.
Ген APC контролирует специфическое сцепление клеток эпителия кишечника, нарушение которого необходимо для возникновения полипа, но недостаточно для возникновения клона раковой опухоли [ Vogelstein, 1988 ]. (Мутация APC ведет к активации бета-катенина , активирующего кадхерин , входящий в состав межклеточных контактов).
Сходная ситуация, по-видимому, имеет место при ХМЛ ( хроническом миелоидном лейкозе ): транслокация BCR-ABL обязательно ведет к ХМЛ, но не сразу после введения филадельфийской хромосомы в клетки человека, трансплантированные бестимусным мышам, а спустя некоторое время и только в немногочисленных клонах клеток [ Daley, 1990 ]. Таким образом, транслокация BCR-ABL резко повышает риск возникновения ХМЛ, но не ведет непосредственно к его возникновению. И в этом случае, скорее всего, появление гена BCR-ABL ведет к предраку , определяющему дальнейшую программу возникновения опухоли.
Суммируя эти ситуации, можно обозначить и второй, чисто генетический путь образования предрака, - формирование компонентов, необходимых для опухоли и входящих в нее, но недостаточных для ее индукции. К этому пути примыкает и весьма распространенный путь инактивации генов-супрессоров опухолей . Ген р53 и родственные ему гены контролируют уход в апоптоз клеток, поврежденных внешними воздействиями, мутациями или старением [ Weinberg, 2006f ]. Мутации этого гена помимо серьезного влияния на регуляцию клеточного цикла индуцируют уход от апоптоза многих клеток, гены которых, проходя кризис, могут приобретать бессмертие и входить в бессмертный генотип опухолевой клетки.
Совершенно неизученной остается проблема эволюции клеток- предшественников опухоли на ранних стадиях, когда они еще не обладают селективными преимуществами, но явно эволюционируют в сторону образования опухолевого клона [ Kern, 1993 ].
Таким образом, многие пути эволюции генотипа образуют типичный предрак , повышая вероятность возникновения определенных опухолей.