Фосфорилаза (гликогенфосфорилаза)
Карта белка гликогенфосфорилазы
G6n+Pi=G6n'+G1P или G6n+Pi=G6ctn'+G1P (n'=n-1)
Катализирует лимитирующую скорость гликогенолиза реакцию: фосфоролитическое расщепление (фосфоролиз) (1->4)-связей гликогена, продуктом является глюкозо-1-фосфат . Остатки глюкозы отщепляются от дальних концов молекулы гликогена ( G6n ) до тех пор, пока на ветвях, идущих от точки ветвления ([1->6]-связи), не останется примерно по 4 остатка глюкозы (такая молекула гликогена обозначена через G6ctn ). n- число остатков глюкозы в молекуле гликогена. См. ( Рис.19.4(БХ) Гликогенез и гликогенолиз: механизм деветвления ).
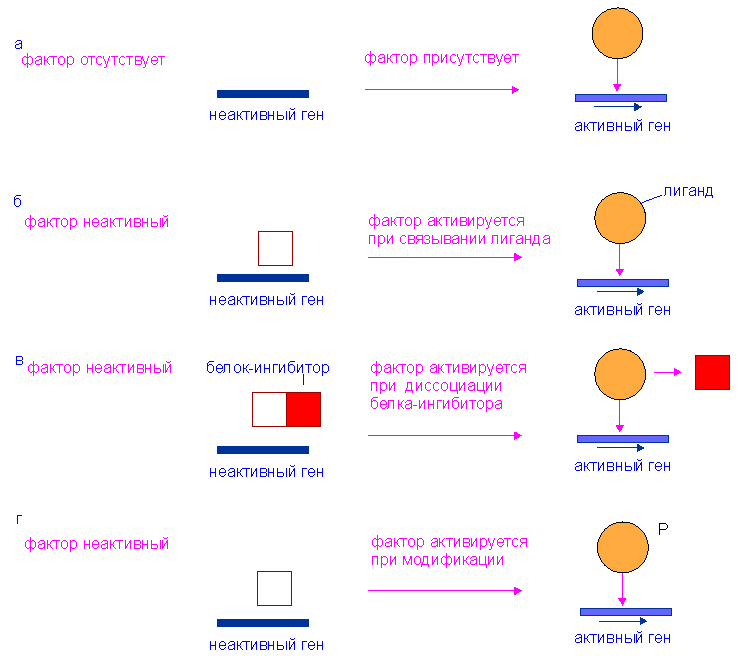
АКТИВАЦИЯ И ИНАКТИВАЦИЯ ФОСФОРИЛАЗЫ ( Рис.19.5(БХ) Механизм: фосфорилаза; регуляция активности )
В печени фосфорилаза находится как в активной, так и в неактивной форме. В активной фосфорилазе (фосфорилазе a) гидроксильная группа одного из остатков серина фосфорилирована. Под действием специфической фосфатазы ( протеинфосфатазы-1 ) фермент превращается в неактивную фосфорилазу b в результате гидролитического отщепления фосфата от остатка серина. Реактивация происходит путем рефосфорилирования за счет ATP при действии специфического фермента киназы фосфорилазы .
Мышечная фосфорилаза иммунологически и генетически отличается от соответствующего фермента печени. Она может находиться в двух формах: в форме фосфорилазы a-фосфорилированного фермента, активного как в присутствии, так и в отсутствие ATP (ее аллостерического модулятора), и в форме фосфорилазы b, дефосфорилированной и активной только в присутствии ATP. Фосфорилаза a является нормальной физиологически активной формой фермента. Она представляет собой димер, каждый мономер которого содержит одну молекулу пиридоксальфосфата.
Активация с участием cAMP
В мышце фосфорилаза активируется адреналином. Однако он оказывает не прямой эффект, а действует опосредованно через cAMP . cAMP представляет собой внутриклеточный интермедиат, выступающий в роли второго посредника при действии ряда гормон ов. Он образуется из ATP при действии фермента аденилатциклазы, находящейся на внутренней поверхности клеточной мембраны. Аденилатциклаза активируется (опосредованно) гормонами адреналином и норадреналином - лигандами (бета-адренергических рецепторов), локализованных в клеточной мембране; в печени она активируется глюкагоном, действующим при участии специального глюкагокового рецептора. cAMP разрушается при действии фосфодиэстеразы ; именно этот фермент поддерживает в норме концентрацию cAMP на низком уровне. Имеются данные о том, что инсулин повышает активность фосфодиэстеразы в печени; это приводит к понижению концентрации cAMP.
Повышение концентрации сАМР активирует фермент, обладающий весьма широкой специфичностью - cAMP-зависимую протеинкиназу. Эта киназа катализирует фосфорилирование при участии ATP неактивной киназы фосфорилазы с образованием активной киназы фосфорилазы, которая в свою очередь, путем фосфорилирования, активирует фосфорилазу b, переходящую в фосфорилазу a.
Фосфорилаза скелетных мышц в отличие от фосфорилазы печени не активируется глюкагоном. Отметим, что фосфорилаза сердечной мышцы активируется этим гормоном, Другим важным отличием является ингибирование печеночной протеинфосфатазы-l активной формой фосфорилазы.
Фосфорилаза a и киназа фосфорилазы- a дефосфорилируются и инактивируются протеинфосфатазой-l. Ингибитором протеинфосфатазы-l является белок, который называют ингибитором-l ; последний становится активным только после фосфорилирования cAMP-зависимой протеинкиназой . Таким образом, cAMP контролирует как активацию, так и инактивацию фосфорилазы.
Гликогенфосфорилаза: литература
Ссылки:
- Инсулин: влияние на утилизацию глюкозы
- Регуляция метаболизма: регуляция метаболизма гликогена
- Ферментный каскад: общая характеристика
- Гликогена метаболизм: введение, биомедицинское значение
- Метаболизм гексоз: фруктоза
- Гликогена метаболизм: механизмы контроля
- Регуляция метаболизма: метаболический контроль ферментативных реакций
- Глюконеогенез: метаболические пути
- Гликогенозы типа III: клиническая картина
- Глюкагон: физиологические эффекты
- Регуляция метаболизма: глюкоза: роль адреналина, глюкагона
- ГЛИКОГЕНОЛИЗ (расщепление гликогена)
- Гликогеноз типа VI (амилопектиноз, болезнь Андерсен (Герса), дефект фосфорилазы в печени)
- Инсулин: влияние на фосфорилирование-дефосфорилирование белков
- Контрактура мышечная