Болезнь Альцгеймера: роль генетических факторов, общие сведения
Прорывом в понимании этиологии болезни Альцгеймера (БА) явилось использование генетических подходов. Интенсивные эпидемиологические исследования позволили выявить родословные, в которых БА характеризуется ранним началом (моложе 65 лет) и моногенным аутосомно-доминантным характером наследования. При более распространенных поздних формах БА (у людей старше 65 лет) также очевидна роль генетических факторов, проявление которых, однако, зависит и от воздействий среды [Pericak-Vance ea 1995 , Rao ea 1995 ]. Генетическая предрасположенность стала четко установленным фактором риска болезни Альцгеймера. Более того, показано, что БА включает в себя несколько генетически гетерогенных форм, объединенных сходными клиническими и гистопатологическими признаками. Причиной или фактором риска развития некоторых (если не всех) форм БА являются мутации или полиморфизмы в ряде генов.
Молекулярные исследования болезни Альцгеймера связывают с идентификацией главного компонента амилоидных бляшек - A-протеина [Glenner ea 1984]. Первый ген и генетический дефект, вызывающий болезнь Альцгеймера, был обнаружен с использованием подхода, который сейчас относят к так называемому "функциональному клонированию". Давно отмечено, что синдром наиболее распространенной врожденной умственной отсталости, синдром Дауна ( с трисомией по 21-й хромосоме), у пациентов старше 35-40 лет также характеризуется гистопатологическими признаками, характерными для БА - у многих больных формируется деменция , накладывающаяся на уже имеющуюся умственную отсталость . Из этого было сделано предположение, что ген (или гены) на 21-й хромосоме могли бы играть решающую роль в нейродегенеративном процессе также и при БА. Действительно, при синдроме Дауна накопление бета-амилоидного белка обусловлено активностью дополнительного гена предшественника бета-амилоидного белка , расположенного на 21-й хромосоме. И в патогенезе по крайней мере некоторых форм болезни Альцгеймера участвуют такие генетические факторы, как ген предшественника бета-амилоидного белка . Кроме того, в единичных семьях с ранней формой семейной болезни Альцгеймера была выявлена точечная мутация в этом гене. Glenner и Wong показали также идентичность амилоидного компонента бляшек при болезни Альцгеймера и при болезни Дауна . Эти случаи послужили первым доказательством возможности моногенного аутосомно-доминантного наследования заболевания (чаще всего мутация приводит к замене валина на изолейцин в положении 717).
К настоящему времени обнаружено, по крайней мере, четыре гена, мутации в которых вызывают БА или являются факторами генетической предрасположенности:
ген белка амилоидного предшественника (АРР) на хромосоме 21 [ Goate ea 1991 ],
ген аполипопротеина Е (АроЕ) на хромосоме 19 [ Sounders ea 1993 ],
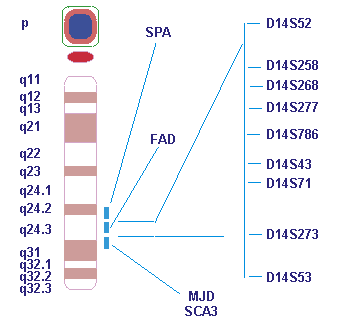
ген пресенилина-1 на хромосоме 14,
ген пресенилина-2 на хромосоме 1 [ Sherrington ea 1995 , Rogaev ea 1995 , Levi-Lahad ea 1995 ].
Один из основных компонентов амилоидных бляшек , которые накапливаются в неокортексе был выделен в конце 70-х годов. Выделенный пептид, названный Ар , как оказалось, состоит всего из 39-43 аминокислотных остатков и является протеолитическим фрагментом более длинного пептида, названного предшественником амилоида ( АРР ) [Kang ea 1987 , Lemaire ea 1989 ].
Ген белка амилоидного предшественника - APP- ген, локализованный на 21-й хромосоме - кодирует белок рецептора наружной клеточной мембраны. Амилоид сенильных бляшек был определен как продукт аномального процессинга APP [ Kang ea 1987 ]. APP-ген стал явным кандидатом для поиска мутаций при болезни Альцгеймера. Действительно, вскоре были обнаружены несколько миссенс-мутаций в экзонах 16, 17, части которых кодируют Ар-пептид [ Goate ea 1991 , Murrel ea 1991 , Chartier-Harlin ea 1991].
Последующий массовый скрининг родословных и спорадических случаев во многих лабораториях мира показал, однако, что мутации в АРР могут объяснить причину БА только 3-5% всех родословных с ранним началом БА.
Дальнейший анализ семейных форм показал, что болезнь Альцгеймера - генетически гетерогенное заболевание. Во многих случаях не наблюдалось сцепленного наследования болезни Альцгеймера с маркерами ДНК 21-й хромосомы [ Hardy ea 2003 , Schellenberg ea 1988 , St George-Hyslop ea 1990 ]. Тем не менее, вскоре была найдена точечная мутация в гене APP, приводившая к развитию болезни Альцгеймера в двух неродственных семьях [ Goate ea 1991 ]. По совокупности биохимических и генетических данных была сформулирована гипотеза амилоидного каскада, в которой первичная роль отводилась отложению нерастворимого амилоида, запускающему дальнейшее развитие болезни Альцгеймера [ Hardy ea 1992 , Hardy ea 2003 ].
В родословных с поздним началом БА мутаций в этом гене не было найдено вовсе [ Tanzi ea 1992 , Schellenherg ea 1991 ].
Впоследствии было установлено, что полиморфизм (эпсилон4-аллель) еще в одном гене, аполипопротеине АроЕ , локализованном на хромосоме 19 , ассоциирован с некоторыми поздними и ранними формами БА [ Sounders ea 1993 , Poirier ea 1993 , Strittmater ea 1993 ]. Такой полиморфизм гена АроЕ, однако, не является необходимым или достаточным условием БА и рассматривается в настоящее время как фактор риска, влияющий на начало развития деменции альцгеймеровского типа [ Bennett ea 1995 ].
Предполагалось, что наиболее вероятными другими генами-кандидатами при БА могли бы стать гены, кодирующие пептиды протеолиза АРР-белка, например, специфические альфа-, Р- или гамма-секретазы; белки, регулирующие синтез АРР; белки нейро-фибриллярных клубков (NFT) или киназы, регулирующие аномальное фосфорилирование отдельных компонентов NFT, например, tau-белков и т.д.
Значительным событием явилось обнаружение семейства генов, ответственных за БА, получивших название гены-пресенилины: ген PS-1 (PSEN1 и ген PS-2 (PSEN2) [Sherrington ea 1995 , Rogaev ea 1995 ]. Важность открытия этих генов обусловлена тем, что были, во-первых, обнаружены принципиально новые, не предполагавшиеся ранее молекулярные компоненты нейропатологии БА, которые были выявлены "слепым" позиционным клонированием, не ориентированным на предыдущие "биохимические" модели болезни; во-вторых, идентифицированы генетические факторы, ответственные за наибольшее число случаев ранних семейных форм БА.. ( Последующий анализ показал, что пресенилины участвуют в процессинге APP, и мутации в генах пресенилинов приводят к увеличению продукции амилоидогенной формы A-бета ( бета-42 пептида ) [ Borchelt, ea 1996 ]).
В начале 90-х годов для поиска новых хромосомных локусов, ответственных за болезнь Альцгеймера, был использован метод классического генетического сцепления с использованием случайных полиморфных маркеров ДНК. В значительной степени такой поиск был ускорен созданием генетических карт хромосом человека и накоплением нового поколения высокоинформативных микросателлитных маркеров (или STR -Simple Tandem Repeats). Ряд таких маркеров, включая STR из хромосомы 14, были выделены к тому времени в лаборатории МГМ НЦПЗ РАМН в рамках проекта Геном Человека , как и в ряде других лабораторий [ Rogaev ea 1992 , Wang ea 1992 ].
Результатом широкомасштабного поиска локуса БА явилось обнаружение в 1992 г. несколькими научными группами участка на хромосоме 14, ответственного за семейные формы БА ( FAD3 ). Было проведено исследование результатов, полученных в Университете г. Торонто при изучении более 20 семей с БА, в которых не было обнаружено сцепления БА с локусом АРР из 21-й хромосомы. После анализа более 70 маркеров из различных хромосом человека максимальный лод-балл и, следовательно, убедительное доказательство существования гена, мутации в котором ответственны за БА, получены для маркера D14S43 (лод-балл больше 20.47) из локуса 14q24.3 (локус FAD3) ( рис. 1 ). При этом было сделано два важных заключения. Во-первых, чрезвычайно высокий суммарный лод-балл, полученный при изучении совокупности семей различного этнического происхождения, свидетельствовал о том, что мутации в гене FAD3 ответственны за многие семейные формы БА (по крайней мере 60-70%) с ранним началом клинического проявления (до 60 лет). Во-вторых, результаты сегрегационного анализа STR-маркеров из локуса 14q24.3 показали, что FAD3 наследуется как моногенное аутосомнодоминантное заболевание, по всей видимости, с высокой пенетрантностью (проявлением мутаций), не зависящей от модификаций генетическими или эпигенетическими факторами.
Генетические факторы, ответственные за наибольшее число случаев ранних семейных форм БА, как было сказано выше, - это гены-пресенилины: ген PS-1 (PSEN1 и ген PS-2 (PSEN2).
Ген PSEN1 расположен на 14-й хромосоме и кодирует белок пресенилин-1 . Мутация этого гена вызывает болезнь Альцгеймера типа 3, которая наследуется аутосомно-доминантно с высокой пенетрантностью. В семьях с самой различной этнической принадлежностью выявлено более 20 мутаций гена PSEN1.
Ген PSEN2 расположен на 1-й хромосоме и кодирует белок пресенилин-2 . Мутация этого гена впервые была обнаружена у американских потомков немцев Поволжья.
Оба гена очень похожи и кодируют сходные белки. Сначала полагали, что эти белки имеют семь трансмембранных доменов, однако дальнейшие исследования показали, что число доменов равно восьми, а девятый располагается у внутренней стороны мембраны. Функция этих белков в норме, а также ее изменения, возникающие в результате мутации кодирующих их генов, неизвестны. Оба белка экспрессируются в нейронах и широко распространены в нервной системе . По первичной структуре они похожи на белок sel12 , участвующий в морфогенезе у нематоды Caenorhabditis elegans .
Мутации генов PSEN1 и PSEN2 сопровождаются повышением концентрации в плазме бета-амилоидного белка , состоящего из 42 аминокислотных остатков, что позволяет предположить взаимосвязь между пресенилинами и предшественником бета-амилоидного белка .
Доказано, что мутации гена PSEN1 - самая частая причина семейной болезни Альцгеймера с ранним началом, обусловливающая 70% случаев этой относительно редкой формы. Болезнь Альцгеймера типа 3, вызванная мутацией гена PSEN1 , характеризуется более ранним началом (в среднем в 45 лет) и более быстрым прогрессированием (средняя продолжительность жизни больного 6-7 лет), чем болезнь Альцгеймера типа 4, вызванная мутацией гена PSEN2 (начало в среднем в 53 года; средняя продолжительность жизни 11 лет). При некоторых редких мутациях гена PSEN2 заболевание начинается после 70 лет.
Роль мутаций этих генов в патогенезе гораздо более частых спорадических форм болезни Альцгеймера с поздним началом не изучена.
В настоящее время разработана методика исследования ДНК клеток крови, позволяющая выявлять такие мутации; пока она применяется только в исследовательских работах. Обследование необходимо при генетическом консультировании лиц с высоким риском заболевания, но без признаков деменции .
Важной вехой стало открытие роли гена, кодирующего апопротеин Е и расположенного на 19-й хромосоме , в развитии поздней семейной и спорадических форм болезни Альцгеймера. Апопротеин Е участвует в транспорте холестерина . Ген апопротеина Е имеет три аллеля: эпсилон2, эпсилонЗ и эпсилон4, кодирующие соответственно изоформы Е2, Е3 и Е4. Аллель эпсилон4 часто обнаруживают при спорадических и поздней семейной формах болезни Альцгеймера. Примерно 24-30% здоровых представителей белой расы имеют хотя бы один аллель эпсилон4, а около 2% - гомозиготны по нему. Среди больных болезнью Альцгеймера от 40 до 65% имеют по крайней мере один аллель эпсилон4 - иными словами, среди больных распространенность этого аллеля существенно выше, чем среди здоровых. С другой стороны, у многих больных аллель эпсилон4 отсутствует и, напротив, имеется у многих здоровых. То есть наличие аллеля эпсилон4 не является ни необходимым, ни достаточным условием болезни Альцгеймера, но служит важным фактором ее риска (особенно у гомозигот). Количество аллелей влияет на возраст начала заболевания: у гомозигот болезнь Альцгеймера начинается раньше. Причины этого не ясны.
Апопротеин Е присутствует в амилоидных бляшках ; он связывается с тау-белком и поэтому, возможно, участвует в образовании нейрофибриллярных включений . Апопротеин Е4 подавляет рост нервных окончаний в культуре клеток спинномозговых ганглиев.
Полагают, что аллель эпсилон2 уменьшает риск болезни Альцгеймера, но эти данные не подтверждены. По непроверенным сведениям, носители аллеля эпсилон4 менее чувствительны к лечению такрином . Есть также данные о том, что аллель A гена альфа1-антихимотрипсина повышает риск болезни Альцгеймера у носителей аллеля эпсилон4.
Единого мнения о целесообразности исследования апопротеина Е в диагностике болезни Альцгеймера нет. У здоровых по результатам этого исследования нельзя судить о риске болезни Альцгеймера, поскольку у многих носителей аллеля эпсилон4 она не развивается. Однако у некоторых гомозиготных носителей аллеля эпсилон4 при позитронно-эмиссионной томографии обнаруживают снижение скорости обмена веществ в коре головного мозга. Некоторые считают эти изменения предвестниками болезни Альцгеймера. Высказывалось предположение, что гомозиготное состояние по аллелю эпсилон4 у пожилого больного с деменцией с вероятностью 96-98% свидетельствует о болезни Альцгеймера. Однако лишь небольшая часть больных гомозиготны по аллелю эпсилон4, и даже у них необходимо исключить обратимые причины деменции. Тем не менее апопротеин Е остается единственным важным биохимическим маркером болезни Альцгеймера.
Продолжается изучение роли апопротеина Е в патогенезе заболевания и его диагностическое значение. Необходимо исследовать взаимосвязь апопротеина Е с другими деменциями.
Показано, что наличие аллеля эпсилон4 не связано с деменцией при болезни Паркинсона и болезни Крейтцфельдта-Якоба .
До сегодняшнего дня APP, PS1 и PS2 остаются единственными генами для которых известны мутации при болезни Альцгеймера. Формы болезни Альцгеймера, вызываемые мутациями в генах APP, PS1, PS2, характеризуются аутосомно-доминантным наследованием, "агрессивным" течением, ранним началом до 65 лет, и составляют до 10% всех случаев заболевания. Все мутации, ассоциированные с болезнь Альцгеймера, суммированы в базах данных по мутациям при болезни Альцгеймера , которые постоянно обновляются (http://www.alzforum.org/res/com/mut/default.asp, http://www.molgen.ua.ac.be/ admutations/).
Интересно, что большая часть найденных мутаций приходится на ген PS1 [ Novak, ea 1993 ], тогда как в APP найдено всего 19 мутаций, а в PS2 - 10. С чем связано такое неравное распределение остается неизвестным.
Помимо классического типа болезни Альцгеймера некоторые мутации в гене PS1 могут также вызывать специфическую форму заболевания, для которой характерны диффузные "ватообразные" сосудистые бляшки, а также симптомы спастического парапареза (неполного паралича конечностей с усиленным тонусом мышц), часто предшествующие деменции [ Crook, ea 1998 ]. Известно, что к такой форме заболевания приводят делеция экзона 10 (по номенклатуре [ Rogaev, ea 1997 ] данный участок соответствует экзону 10), а также около 10 других миссенс-мутаций в PS1 [ Hattori, ea 2004 ]. Механизм действия мутаций, приводящий к данной патологии, до сих пор не изучен.
Второй главный маркер болезни Альцгеймера - нейрофибриллярные клубки - также интенсивно изучается. Найдено, что нейрофибриллярные клубки состоят в основном из конгломератов гиперфосфорилированной полной формы тау-белка , а также из фрагментов тау-белка [ Grundke-Iqbal, ea 1986 , Grundke-Iqbal, ea 1986 , Novak, ea 1993 , Wood, ea 1986 ]. Был клонирован тау-ген ( MAPT, Microtubule-Associated Protein Tau ген ), который локализован на 17-й хромосоме [ Neve, ea 1986 ]. Мутаций, приводящих к болезни Альцгеймера, в MAPT гене обнаружено не было. Интересно, что в недавних исследованиях в MAPT гене было найдено более 30 мутаций, вызывающих некоторые формы лобно-височной деменции , при которой в мозге пациентов образуются нейрофибриллярные клубки, такие же как при болезни Альцгеймера, но отсутствуют отложения b-амилоида [ Forman, ea 2004 ]. В гипотезу амилоидного каскада было внесено уточнение: при болезни Альцгеймера нерастворимый амилоид запускает процесс, который через нарушение структуры микротрубочек приводит, в конечном итоге, к гибели клеток и нейродегенерации. При лобно-височной деменции непосредственно мутации в MAPT гене являются причиной образования нейрофибриллярных клубков [ Hardy, ea 1998 ].
Найдены три мутации в гене пресенилина 1, ассоциированные с развитием лобно-височной деменции, но не болезни Альцгеймера. Предполагается, что такие мутации в PS1, могут приводить к частичной потере функций гена, и как следствие - отсутствию накоплений амилоида [ Amtul, ea 2002 , Dermaut, ea 2004 , Raux, ea 2000 ].
Хотя эти данные неоднозначны и требуют дальнейшего исследования, не исключается существование "неамилоидных" механизмов развития деменции при мутациях в гене PS1.
{kind=link}